During my sweeping tweaks to my carcharodontosaurid reconstructions I decided to do a volumetric estimate for my updated reconstruction of Giganotosaurus carolinii.
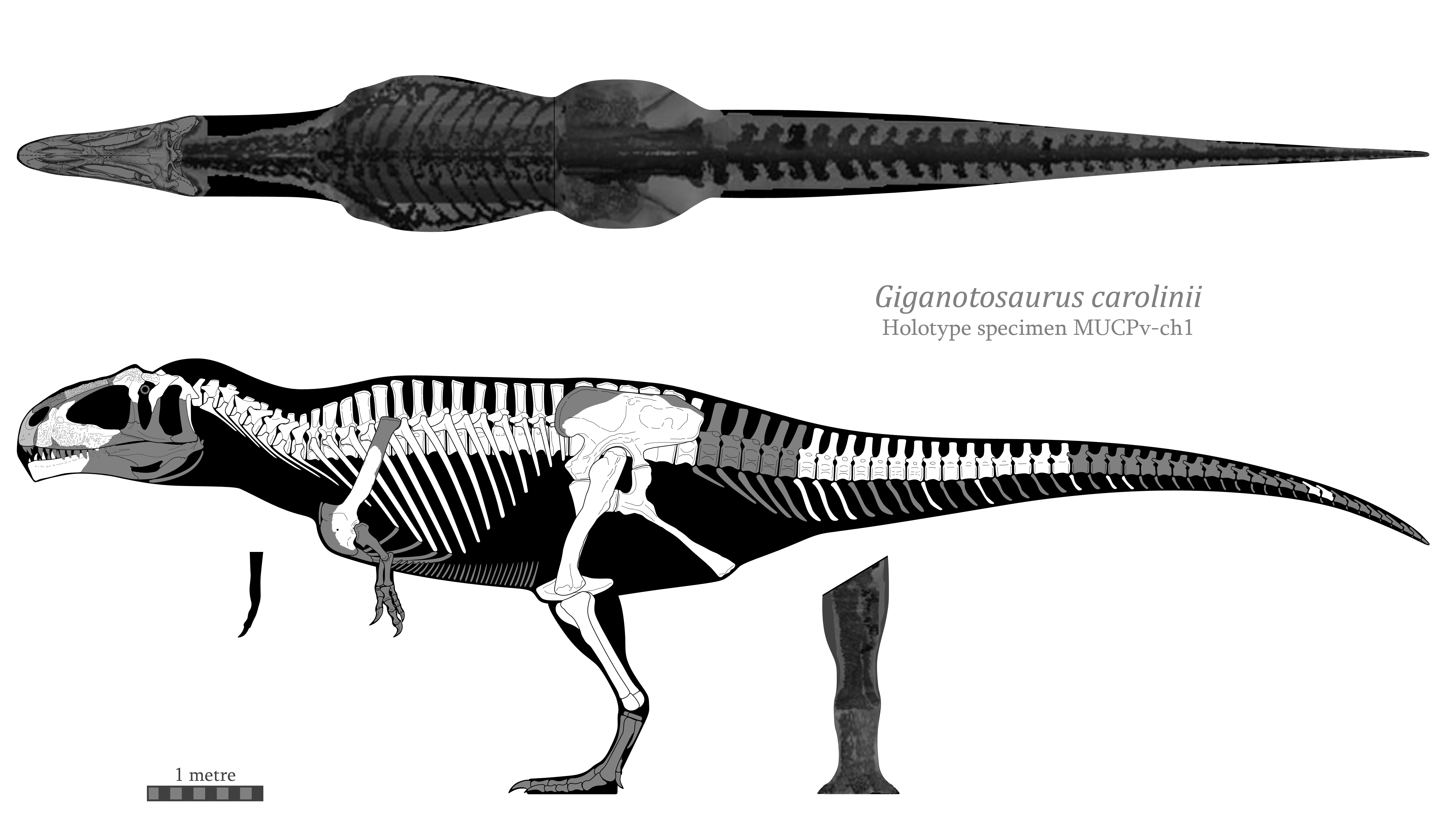
For the dorsal view and the anterior views of the limbs, I took the scanned Acrocanthosaurus skeleton from Bates et al. (2009), scaled and modified to fit the Giganotosaurus. I then took the dorsal view of the Carcharodontosaurus skull from Sereno (1996) and adjusted it to fit.
Using a Python script to perform a graphic double integration with one-pixel slices at a scale of 364 px = 1 metre, the estimated volumes for each section were as follows:
| Section | Volume (L) |
| Head | 497 |
| Neck | 593.5 |
| Torso | 5281.4 |
| Tail | 1621.2 |
| Individual forelimb | 7.2 |
| Individual hindlimb | 296.7 |
| Total | 8601 |
Sections can be downloaded here.
The estimated mass for MUCPv-ch1 using a density of 0.95 kg/L (Larramendi, Paul, & Hsu, 2020) would be around ~8.17 tonnes.
Readers might note that this is significantly larger than the 2013 mass estimate by Scott Hartman of 6.8 tonnes for the same specimen. This might seem egregious especially given how my own reconstruction bases several aspects on it due to the dearth of available data. Given how touchy people can get about megatheropod mass estimates, this might generate some less-than-friendly dissent thrown my way, but let me explain. There are a few reasons for this.
For one, the 2013 Hartman Giganotosaurus estimate has an average bodily density of about ~0.913 kg/L. With a density of 0.95 kg/L, it would raise the resulting mass to about ~7.1 tonnes even when the above factors were disregarded. Furthermore, there’s quite a precision level difference here that contributes to the disparity. The 2013 Hartman GDI used 46 equally spaced slices along the axial body, a few slices for the hindlimbs and 10 for the hindlimbs, rather than a pixel-precise one as done here.
Those differences by no means account for the large disparity in the final result – they aren’t close to being big enough on their own. By far, the biggest factor is the restoration of the shoulder girdle and its effect on the chest.
In Scott Hartman’s reconstruction the Giganotosaurus type scapulacoracoid (preserved length ~90 cm) was taken to be rather well-preserved, assuming that only a small portion of the distal end of the scapula was missing, and restores the total scapulacoracoid length at ~97 cm. This results in the diminutive pectoral girdle the animal had been traditionally reconstructed with.
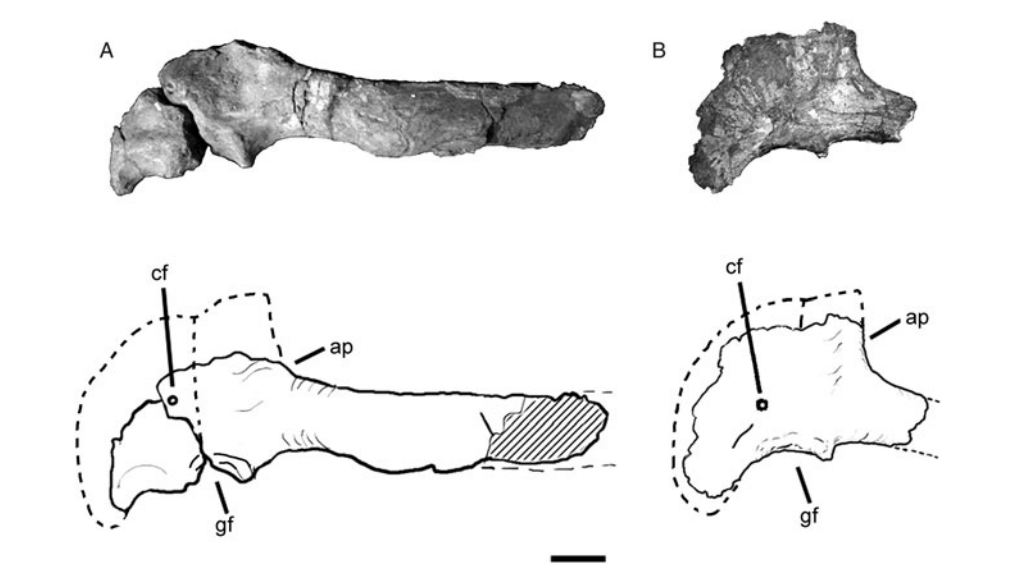
However, the scapulacoracoid in actuality is very badly and incompletely preserved, with the acromial process and a major portion of the coracoid broken off, which lead to misinterpretation of its anatomy (Canale et al. 2013). Furthermore, comparisons with other carcharodontosaurids also indicate a significantly longer scapulacoracoid. I have estimated the complete length of the scapula alone at ~103.7 cm based on Acrocanthosaurus, assuming a 127.7 cm long femur for NCSM 14345 (Carpenter 2000) and the scapula:femur proportions being similar between the two taxa. This would be a minimalistic estimate, as the femur of NCSM 14345 is incompletely preserved and the aforementioned length being probably overinflated – comparisons with the complete femur of SMU 74646 yields an est. length of ~119.4-123.1 cm, which would consequently increase the estimated scapula length for Giganotosaurus. Nevertheless, the minimalistic estimate was used here. Adding in the coracoid gives a total length of ~125 cm.
The resulting larger pectoral girdle deepens the anterior chest significantly. With the estimated width here being identical (134 cm including soft tissues) the chest ends up considerably larger, and since it is the most voluminous portion of the animal’s body and comprises a major portion of the total, the overall volume and mass of the animal shoots up considerably. The hips of my reconstruction are a bit shallower, but due to the lower volume of the pelvic region it is nowhere near enough to counteract the aforementioned increase.
I have no guarantees that everyone will be satisfied by this explanation, especially those among the many palaeoenthusiast communities all over the internet that take theropod mass estimates way too seriously, but it should serve to explain why my estimate is so much higher. But I digress.
If this estimate holds, then Giganotosaurus would truly deserve its name. For comparison, average African elephants mass around ~4 and ~6 tonnes for adult females and males respectively (Larramendi 2016). The chances of the single good specimen of Giganotosaurus we have being the largest that ever lived is also vanishingly small, and statistically speaking it would most likely have been among the average-sized ones as there are much more of those in any animal population than there are especially large or small individuals. There might have been some real monster giganotosaurs out there for all we know!
This also has implications on a rather…fringe hypothesis made recently: that allosauroids were supposedly obligate scavengers akin to vultures as proposed by Pahl & Ruedas (2021). While they used a 2-tonne Allosaurus as their model theropod, Giganotosaurus as an allosauroid would have also fallen under this umbrella. The hypothesis is hugely flawed in several aspects, but the relevant one to this post here pertains to the relationship between body size and the energy efficiency of a scavenger lifestyle.
A study by Kane et al. (2016) models theropods of various sizes foraging and competing for carrion to estimate the energy expenditures and gains from scavenging. According to their model the most efficient size range for scavenging in theropods was estimated at 27-1044 kg.

At body masses over a tonne, energy intake from scavenging plateaued due to a limited accessible supply of carcasses after competition. While gigantic megatheropods had much larger gut capacities and capability to take over carcasses from smaller animals, the costs of foraging outpaces the energy gain from scavenging at massive sizes. The 2-tonne Allosaurus used by Pahl & Ruedas for their model is already about twice as massive as the upper reaches of the optimal scavenging range, putting severe aspersions to their hypothesis. At over 8 tonnes, the Giganotosaurus estimated here is much too massive for scavenging to be a remotely sustainable lifestyle beyond opportunistic occasions. Forced into an obligate scavenger lifestyle, it would fail to meet its energy demands by far and starve to death. Theropods of this size are thus very unlikely to have been driven by scavenging and most likely were specialised hyperpredators.

Acknowledgements
Credit to Franoys for the Python GDI script.
References
- Sereno et al., 1996, “Predatory dinosaurs from the Sahara and Late Cretaceous faunal differentiation”
- Carpenter, 2000, “A new specimen of Acrocanthosaurus atokensis (Theropoda, Dinosauria) from the Lower Cretaceous Antlers Formation (Lower Cretaceous, Aptian) of Oklahoma, USA”
- Bates et al., 2009, “Estimating mass properties of dinosaurs using laser imaging and 3D computer modelling”
- Canale et al, 2013, “Osteology and phylogenetic relationships of Tyrannotitan chubutensis Novas, de Valais, Vickers-Rich and Rich, 2005 (Theropoda: Carcharodontosauridae) from the Lower
- Kane et al. (2016), “Body size as a driver of scavenging in theropod dinosaurs”
- Larramendi, 2016, “Shoulder height, body mass, and shape of proboscideans”
- Larramendi, Paul, & Hsu, 2020, “A review and reappraisal of the specific gravities of present and past multicellular organisms, with an emphasis on tetrapods”
- Pahl & Ruedas, 2021, “Carnosaurs as apex scavengers: Agent-based simulations reveal possible vulture analogues in Late Jurassic dinosaurs”

Seeing as how I got 7-8 tonnes in Princeton Field Guide to Dinosaurs 2nd edition with a lower SG can’t complain about this result. Gregory Paul
LikeLike
Awesome work.
Im looking forward to seeing your T Rex and spinosaurus mass estimation post.
LikeLike
Finally, SOMEONE acknowledges that scavenging theropod paper. Giant theropods must have had an interesting biology in life, given all the hunting they would have had to rely on. Also makes you wonder how horrible the K-Pg event was once most of the live megafauna were dead and the carnivores had to resort to scavenging (or preying on other carnivores).
LikeLike
Actually, forget the extrapolations, what I got for the distal limb elements based on the difference between hartman’s volumes and mine is way too high. I now checked and am getting 382 l for both distal hindlimbs of sue, which is a bit on the low side but way more consistent with other analyses than the enormour 1000 l from my last comment. So I think I am getting somewhat lower values than Hartman using the pixel-precise slicing, with the overall volume ending up around 8600 l rather than the 9200 he got. The same, presumably, would hold true for Giganotosaurus.
LikeLike
Late reply, my apologies but I’m not as active as I used to be.
I’m unable to approve your previous comment so it can show up, did you delete it? Well, anyway I still have it in my email inbox so I can still reply to it. Adding the fore- and hindlimbs of this model to the axial body of yours, I get ~8505 L (~8080 kg using an SG of 0.95). But the belly of the lateral you used (https://sta.sh/021aolq1ymib) seems punched in by quite a bit XD. Considering that, and differences in neck tissue, skull reconstruction, and other uncertain variables, it’s pretty close. As for Sue, adding your 382 L hindlimbs to the 9294 L axial body your provided gives me 10,058 L (~9555 kg at 0.95 SG). Messed up something in the math perhaps?
As for MUCPv-95, while I ignored it in this post, it most likely rivals Sue. I’m in a Discord server with someone who had personally gone and measured bones of the Giganotosaurus holotype, and the dentary of MUCPv-95 scales some ~6.6% longer than that of MUCPv-ch1 measured to the 13th alveolus. You can try to contact Darknix (that someone, if you see “Sanctum Archaic Stories” along with that name then you got the guy) for more info that he is at liberty to share.
As for Scotty, it might well be similar in size to Sue, or perhaps smaller if the torso length I’ve got from scaling laterals of its mounts is correct XD. It’s femora, hips, etc. are bigger sure, but a shorter trunk would do a lot to offset it.
I have a new reconstruction of Giganotosaurus in the works as I had received both new and previously-overlooked data since then, as well as re-evaluating some previous data sources I used for the old version, although it’s proceeding slowly due to my motivation to work on palaeo content going in and out like a tide. But I will say that it’s actually likely even more massive than the version here mainly due to a longer sacrum, deeper hips, and a bigger tail. To elaborate further on those three major proportional differences respectively:
1. The supplementary data of the Murusraptor description (Coria & Currie 2016) has a measurement of the holotype’s sacrum at 109.2 cm, while this version and Hartman’s has it at ~90 cm. This of course makes the torso a bit longer.
2. This version uses measurements taken from the Theropod Database (111 cm pubis and 120 cm ischium). However after thinking about it some time and some scaling attempts, as well as looking at it with a new lens after the publication of the 2022 Meraxes description, I have arrived to the conclusion that these measurements were very likely erroneous and probably taken from the…weird skeletal reconstruction…thing…from the original 1995 description. When I try to scale the mounts’ hips based on reported measurements of the femur, the pubis always ends up significantly longer than the TD’s figure (the ischium on the other hand does come pretty close), and furthermore the proportions seen in Meraxes puts extra aspersions on the idea of Giganotosaurus having a shorter pubis than its ischium. The TD entry on Giganotosaurus also lists 154 cm for the ilium, which I’m sure is way off.
3. Molina-Perez & Larramendi’s book on theropods (2016, 2019 for the English translation) has a measurement for the longest chevron in the Giganotosaurus holotype. It’s 48 cm, a bit over 13% longer than the longest one in Hartman’s.
LikeLike
Hi!
Not sure what happened to my first comment, but I commented as guest, so I can’t delete anything (otherwise I actually would have deleted my second comment, which I added thinking it would also need approval before appearing, so that both comments would turn up at the same time lol).
Well anyway, I think I found the issue with my previous figures; the scale bar in the original file is 98 px rather than 100 px, but I just assumed back then that that’s what it was supposed to be, but I now re-measured and checked sue’s axial length and other dimensions, which end up the correct size when the whole thing is scaled up to 100 px scalebar length. With the rescaled dimensions. I also think I didn’t do the hindlimbs well last time (I sort of rotated and stretched the silhouettes into allignment rather than just adding a bit of the thigh on top of the part that protrudes from the silhouette of the trunk.
Corrected results: Axial segment 8798.959 l
dist. hindlimb 238.4712, ×2= 476.9425 l
Total: 9275.902 l
So I am very close to Hartman’s original figure. (I’m also getting very similar figures for Giganotosaurus, at a total volume of 7437.5140l).
So basically, forget my last comment lol
Looking forward to your new Giganotosaurus reconstruction. It really still boggles the mind how one needs to piece together the information on such a relatively complete specimen known for 30 years from little mentions in appendices and supplements. Really hope someone at some point is going to give it a proper description.
LikeLike
what are you talking about?
LikeLike